
| Biyoloji alt dalı |
| Evrimsel biyoloji |
|---|
 |
|
Evrimsel biyoloji portalı Kategori • İlgili başlıklar • Kitap |

İnsanın evrimi, modern insanın (Latince: Homo sapiens) evrimsel kökenini ve ne tür evrimsel süreçlerle ortaya çıktığını incelediği gibi insanın en eski atalarını ve atasal kökenlerini de konu edinir. Bunun yanında insanla ortak ataları paylaşan ve insan ile yakın akraba olan türlerin evrimini ve kökenini de araştırır. İnsan evrimi, konu olarak 1863 yılında T. H. Huxley tarafından oluşturulan bilim dalı primatolojiyi ve günümüz maymunlarının tüm yaşam formları ile onların eski atalarının fosillerini de dikkate almaktadır. Bunun yanında insanın evrimsel tarihi üzerindeki çalışma ve araştırmalar fiziksel antropoloji, paleoantropoloji, primatoloji, arkeoloji, dilbilim, genetik ve embriyoloji dahil olmak üzere birçok bilimsel disiplinleri de içerir.
Genetik araştırmalar ve fosil kayıtlar ışığında, insanın da üyesi olduğu Primat takımının evrim sürecinde günümüzden yaklaşık 65-55 milyon yıl öncesinde Paleosen dönemde ortaya çıktığı tespit edilmiştir.Hominidae ailesi veya büyük insansı maymunlar, 15-20 milyon yıl önce Miyosen dönemde Hylobatidae (Gibongiller) ailesinden ayrılmıştır. Yaklaşık 13 milyon yıl önce Ponginae veya orangutanlar Hominidae ailesinden ayrılmıştır. Goril ata formlarının insan ve şempanze'nin dahil olduğu Hominini oymağını oluşturan türlerden 8,5 ila 12 milyon yıl önce ayrıldığı tahmin edilirken, insan ile şempanze cinslerini oluşturacak ataların iki alt oymağa (Hominina ve Panina) 7 ila 5,5 milyon yıl önce ayrıldığı düşünülmektedir.Homo cinsi bundan yaklaşık 2,5 milyon yıl önce Afrika'da hominin türlerinden evrilmiştir. Bu anlamda insanın yakın dönem evrimi, insan ve şempanzelerin ortak ataları olan toplulukların birbirinden ayrılmasından sonra başlamıştır. Bu iki topluluktan insanın ortaya çıktığı grup ile bu grubun tüm soyu tükenen türleri hominin olarak adlandırılırlar.
Afrika, Asya ve Avrupa'nın farklı bölgelerinde giderek artan sayıda iyi korunmuş fosillerin bulunuşu, insan evrimi hakkındaki bulguların genişlemesine ve insanların ataları hakkında bilgilerin artmasına neden olmakla birlikte bu bulgulardan elde edilen bilgiler, insan ve insan öncesi türlerin tam olarak yaşadıkları zaman aralıkları ile dağılım bölgelerinin eksiksiz olarak belirlenmesinde ve bilim insanları arasında tam bir görüş birliği sağlanmasında yeterli olmayıp buna dair daha fazla tartışmaların ve kapsamlı araştırmaların yapılmasını gerekli kılmaktadır. İnsan evrimi hakkındaki araştırmalar, mevcut yöntemlerin sınırlarında ya da bu bulguların sağladığı analitik verilerin altında hareket ettiği için insanın soyağacı hakkındaki tartışmalar, insanın atası olan tarih öncesi birçok türün soyağacındaki yerlerinin sistematik sınıflandırması ile bu türlerin birbirleriyle olan akrabalık dereceleri ve yakınlıkları tartışmalı olup tam olarak tamamlanmamıştır.
Çoğu kez Hominini türleri şu şekilde belirlenmiştir: İnsandan önce insana yakın türleri içeren ve "ön insan" denilen australopitesinler; "ilk insanlar" olarak tanımlanan Homo habilis ve Homo rudolfensis; "erken insanlar" olarak daha sonra ortaya çıkan ve Homo cinsine dahil olan tüm türler (Homo sapiens hariç); "modern insan" veya "günümüz insanı" olarak tanımlanan Homo sapiens. Bunun yanında insanın evrimine, bazen insandan önceki bir zaman dilimi olan ve günümüzden 23 ila 5,3 milyon yıl öncesini kapsayan Miyosen dönemindeki hayvandan insana geçiş aşaması da dahil edilir.
İnsanın oluşumu
Bir hücreden insana:

4.57 milyar yıl önce oluşan Yer üzerindeki yaşamla ilgili ilk kesin kanıtlar, 3.5 milyar yıl öncesine (Arkeen) tarihlenir. Moleküler genetik çalışmalar, bunu (LUCA) 4.4 milyar yıl öncesine kadar geri çeker.
Arkeen'de en eski bakteriler ve arkeler ortaya çıkmıştır. Genetik çözümlemelere göre ökaryotlar, arkelere kalıtsal olarak daha yakındır. 1980'lerde James Lake tarafından önerilen eosit hipotezi, ökaryotların prokaryotik eositler adında bir arke grubunun içinde ortaya çıktığını öne sürer.
Crenarchaea ve ökaryotlar arasında yakın bir ilişkiyi destekleyen olası bir kanıt, Crenarchea'da RNA polimeraz alt birimi Rbp-8'in bir homologunun varlığıdır, ancak bu Euryarchaeota'da bulunmaz (bkz. Eosit hipotezi).
2017'de yayımlanan bir çalışmada, bir Proteoarchaeota grubu arke (Asgardarchaeota), ökaryotların yan taksonu olarak sınıflandırılmıştır.
Endosimbiyoz teorisine göre alfaproteobakteriler ile ortakyaşamlı olmuş bir arke, ökaryot üst alemini oluşturmuştur. Bu alfaproteobakteri mitokondriye dönüşmüştür. Ayrıca ökaryot bir hücrenin içinde yaşamaya başlayan bir siyanobakteri, bitkiler ve yosunlarda bulunan plastidlere dönüşmüştür. Fosil kayıtlarında en eski ökaryotlara, 1.6-1.7 milyar yıl öncesinde rastlanır.

İnsanın içinde bulunduğu hayvanlar aleminin kalıtsal olarak en yakın akrabası yakalı kamçılılardır. Hayvanların atalarının onlara benzemiş olabileceği düşünülmektedir. Yakalı kamçılılar tek hücrelilerdir. Grubun, hayvanlar ile son ortak atasının yaklaşık 900 milyon yıl önce yaşadığı düşünülmektedir. "... [yakalı kamçılılar] süngerin su kanallarının yüzeyini kaplayan koanositlere çok benziyorlar. Uzun süre ya sünger bir atanın bir kalıntısını temsil ettiklerinden ya süngerlerin bozulup tek ya da çok az hücreye dönüşmüş evrimsel torunları olduklarından kuşkulanıldı. Moleküler genetik kanıtlar ilkini gösteriyor..."


En eski hayvan fosillerine Ediyakaran (635-538.8 myö) döneminde rastlanır. Ediyakaran, sanılana göre hayvanların çok hücreli atalarının ortaya çıktığı dönemdir. Bu dönemin canlılarına Ediyakaran biyotası denir. Yaklaşık 538.8 milyon yıl önce gerçekleşen Kambriyen patlaması ile günümüz hayvan şubelerinin ilk üyeleri ortaya çıkmıştır. İnsan, hayvan şubeleri arasında, daha çok omurgalıları içeren kordalılardandır. Kordalıların son ortak atasının yaklaşık 560 milyon yıl önce yaşadığı düşünülür. Tüm omurgalıların son ortak atası ise sanılana göre yaklaşık 530 milyon yıl önce yaşamıştır. "... [tüm omurgalıların son ortak atasının] neye benzediğini kimse bilmiyor; ama muhtemelen Devoniyen Balık Çağı'ndan ve korkunç denizakreplerinden çok önce, Kambriyen Dönem'de yaşadığına göre, çenesiz balıkların altın çağının zırhlı balıkları gibi zırh-levhalı değildi. Yine de, zırhlı balıklar taşemenlerden çok, biz çeneli omurgalıların yakın kuzenleri gibi görünüyorlar."

İnsanın da bulunduğu kemikli balıklar ile köpekbalığı, vatoz gibi omurgalıları içeren kıkırdaklı balıkların son ortak atası muhtemelen Ordovisyen dönemde (460 myö) yaşadı. Kemikli balıklar iki gruba ayrılır: et yüzgeçliler ve ışınsal yüzgeçliler. İnsanın bulunduğu et yüzgeçliler ile yüzme keseleriyle tanınan ışınsal yüzgeçlilerin son ortak atası, sanılana göre yaklaşık 440 milyon yıl önce Silüriyen dönemde yaşamıştır.

Devoniyen dönemin ortalarında (390 myö) karaya çıkmaya başlayan et yüzgeçliler Tetrapodomorpha adlı yeni bir soy oluşturmuşlardır. Düşünülene göre akciğerler ilk olarak tüm kemikli balıkların son ortak atası tarafından geliştirilmiş ancak bu akciğerler ışınsal yüzgeçlilerde yüzme keselerine dönüşmüştür (bkz. Akciğerin evrimi). Dört üyeliler tetrapodomorfların taç grubudur ve tüm dört üyelilerin son ortak atası (bir iki yaşamlı ile insanın son ortak atası) yaklaşık 340 milyon yıl önce yaşamıştır.

İkiyaşamlı olmayan tüm günümüz dört üyelileri amniyottur ve son ortak ataları karaya yumurtlama becerisini geliştirmiştir. Amniyotların ataları bundan 310 milyon yıl önce ayrılmış ve Sauropsida ve Synapsida adlı iki soyu oluşturmuşlardır. Sauropsida, kuşları ve sürüngenleri, Synapsida ise memelileri barındırır.
İlk olarak Geç Permiyen döneminde (260 myö) ortaya çıkmış sinodontlar memelilere yol açmıştır. En eski kılların sinodontlar tarafından geliştirildiği düşünülüyor. Kıllara dair en eski kanıtlar, erken Jura dönemine dayanır (bkz. Morganucodon).


En eski memeliler, fosil kayıtlarında yaklaşık 210 milyon yıl önce ortaya çıkmıştır. Günümüz memelilerinin son ortak atasının (ornitorenk ya da ekidne ile insanın son ortak atası) yaklaşık 180 milyon yıl önce yaşadığı düşünülmektedir.Doğurgan memelilerin çesitlenmesi (bir keseli ile insanın son ortak atası) ise yaklaşık 140 ila 130 milyon yıl önce gerçekleşmişir.

En eski eteneli memelilerin yaklaşık 105 ila 120 milyon yıl önce 2 ayrı gruba ayrıldığı düşünülüyor: Afrotheria ve Xenarthra'yı kapsayan Atlantogenata ve insanın dahil olduğu Boreoeutheria. (Bir diğer varsayım Afrotheria'yı Boreoeutheria'nın kardeş taksonu olarak sınıflandırır.)
Moleküler genetik çalışmalara göre, Boreoeutheria'nın yaklaşık 95 ila 100 milyon yıl önce, insanın dahil olduğu Euarchontoglires ve Laurasiatheria adlı iki gruba ayrılmıştır.


Primatların ortaya çıkışı:
- 86-90 milyon yıl önce, Glires (kemirgenler ve tavşanımsılar) grubu ile euarchontanların atalarının iki ayrı soy hattı oluşturması.
- 80-86 milyon yıl önce, sivri sincapçıklar (scandentia) ve primatomorfların atalarının ayrışması.
- 80 milyon yıl önce, abalı memeliler ve primatların atalarının iki ayrı soy hattı oluşturması.
- 80-63.3 milyon yıl önce, başparmağı öteki parmaklarının karşısına konuşlanabilen canlıların (gerçek primatlar) yeryüzünde görülmeye başlanması.
- 55 milyon yıl önce göz çukurları öne kaymış olup üç boyutlu görebilen primatların (önmaymun) ortaya çıkması.
- 40 milyon yıl önce ilk simiyenlerin ortaya çıkışı.
- ~33-40 milyon yıl önce simiyenlerin iki parvo takıma ayrılıp Yeni Dünya maymunları ve Eski Dünya maymunlarının görünmeye başlaması.
- 23-33 milyon yıl önce; insansı maymunların diğer köpeksi maymunların (Cercopithecoidea) atalarından ayrılması.
Fosil kayıtlarında yaklaşık 63 milyon yıl önce ortaya çıkan primat takımının 60-75 milyon yıl önce Strepsirrhini ve Haplorrhini adlı iki primat alt takımına ayrıldığı düşünülmektedir, ancak fosil kaydına göre en eski önmaymun fosili 55 milyon yıl öncesine tarihlenmiştir.
Primatların filogenetik ağacı
|
Hominoidea (İnsansılar)

Büyük insansı maymunlar ve gibongillerin son ortak atası 16 ila 20 milyon yıl önce yaşamıştır.
- Hylobatidae - (Gibongiller)
- Hominidae - insan türüne giden yol
Hominid (Büyük insansı maymunlar)
Hominidae, tarih öncesi insansı canlıları ve yaşayan Pan (şempanze), Gorilla (goril), Pongo (orangutan) ve Homo (insan) cinsini kapsayan bilimsel familyaya verilen addır.
Büyük insansı maymunlardan soyu orangutana varacak olan Ponginae familyası yaklaşık 12 ila 16 milyon yıl önce ayrılmıştır.
Homininae
Yaklaşık 8.5 ila 12 milyon yıl önce günümüzün Gorilla, Pan ve Homo cinslerini oluşturan atalar birbirinden ayrılmıştır.
Hominini (İnsansılar)
Şempanzelerin ve insanların son ortak atası, günümüzden 5.5 ila 7 milyon yıl önce birbirinden yaşamıştır.

Hominina (İki ayak üzerinde durabilen insansılar) Bipedalizm
- Sahelanthropus, yaklaşık 7 milyon yıl önce yaşadı.
- Orrorin, yaklaşık 6 milyon yıl önce yaşadı.
- Ardipithecus, 5,6 milyon yıl önce ortaya çıktı ve 4.4 milyon yıl önce soyu tükendi.
- Australopithecus, 4,2 milyon yıl önce evrimleşti - 1,3 milyon yıl önce soyu tükendi.
- Kenyanthropus, 3,5 milyon yıl önce yaşadı.
- Paranthropus, 2.7 milyon yıl önce evrimleşti, 1,3 milyon yıl önce -0.8 milyon yıl öncesine ait olası kalıntıları da bulunur- soyu tükendi.
Homo (insan ve yakın akrabaları)
- Homo habilis (yetenekli insan), bilinen en eski Homo türü, 2.4-2.3 milyon yıl önce ortaya çıktı.
- Homo rudolfensis, 2.4 milyon yıl önce ortaya çıktı. Kimilerine göre Homo habilis ile eş anlamlı.
- Homo ergaster (çalışan insan), ~ 2 milyon yıl önce ortaya çıktı. Kimilerine göre Homo erectus ile eş anlamlı.
- Homo gautengensis, 1.9-1.8 milyon yıl önce evrimleşti.
- Homo erectus (dik insan), Homo habilis'ten türemiş ve daha gelişmiş insan atası, 2 milyon yıl önce evrimleşti, 110 000 yıl önce Cava adasında yok oldu.
- Homo georgicus (Gürcistan insanı), 1.8 milyon yıl önce evrimleşti.
- Homo cepranensis, 800 000 yıl önce yaşadı.
- Homo antecessor, 900 000 yıl önce yaşadı.
- Homo heidelbergensis, 600 000 yıl önce evrimleşti "Goliath" (Hem neandertallerin, hem de modern insanın en olası ortak atası olarak kabul edilir.).
- Homo neanderthalensis, (Neandertal insanı) ~ 300 000 yıl önce evrimleşti - 40 000 yıl önce yok oldu.
- Homo rhodesiensis, 800.000 yıl önce evrimleşti, 120.000 yıl önce yok oldu
- Denisova insanı, Altay bölgesinde bulunan fosilleri 40 000 yıl öncesine tarihlendi.
- Homo floresiensis (Flores insanı), 190 000 yıl önce evrimleşti, 40 000 yıl önce soyu tükendi ("Hobbit").
- Homo sapiens, bilge insan, 300 000 - 350 000 yıl önce ortaya çıktı.
- Homo sapiens idaltu, 160 000 yıl önce yaşadı.
- Homo sapiens sapiens, modern insan, son ortak atası ~200.000 yıl önce yaşadı.
İnsansıların kökenine dair molekülergenetik bulgular
Fosiller aracılığıyla türler tek tek birbirlerinden ayırt edilebildiği gibi var oldukları da bu fosiller temelinde belgelenip kanıtlanabilmektedir. Buna karşın belirli bir türün bireylerinin fiziksel özelliklerinin onları daha genç ve erken bir döneme atacak kadar evrimleşip farklılaştığı tam zaman noktası fosiller temelinde sadece kaba bir şekilde tahmin edilebilirler. Bunun nedeni, bir önceki türün ilk özellikleri ile bir sonraki türün yeni kazanılmış özellikleri arasında keskin olmayan, tam aksine çok yumuşak ve akıcı geçişlerin bulunmasıdır. Birbirini takip eden iki tür arasında keskin bir çizgi çizmek zor olduğu gibi çoğu zaman fosil bulgulardaki eksiklikler ve aralıklar yüzünden de neredeyse imkânsızdır.
Moleküler saat, türlerin birbirinden ayrıldığı zamanı daha tam ve daha yakın olarak tespit etmek için araştırmalarda kullanılan önemli bir metot olup bunun için günümüzde yaşayan türlerin DNA analizlerine ihtiyaç duyulur. Moleküler saatin tam akış hızı ve geçmiş dönemlerde mutasyon görülme sıklıkların oranı tam olarak bilinmediği ve kalibrasyon karşılaştırmaları fosil kalıntılara bağlı olduğu için bu değerlendirmelerin sonuçları uzmanlar arasında tartışmalı olup sadece primat evrimin erken aşamaları için de söz konusu değildir. Bu anlamda 1960'ların sonlarında primatların evrimi için oluşturulan zaman çizelgesi ilke olarak hala geçerliğini korusa da detaylara girildiğinde fazlaca açıldığını ve yayıldığını göstermiştir. Bunun yanında moleküler saat ile hesaplanan zamanlar ile fosil bulguların tarihlendirilmeleri ile elde edilinen zamanlar arasında çoğu zaman milyonlarca yıl fark bulunabilmektedir.

Bu anlamda, 1985 yılında DNA analizlerin sonucuna dayalı olarak yayınlanan bir çalışmaya göre 90 milyon yıl önce Euarchontoglires ailesinden diğerlerinin yanında bir taraftan günümüz fareleri ile diğer taraftan içinde büyük insansı maymunlar ve gibongillerin de yer aldığı insansılar oluşmuştur. Son analizler de aynı şekilde bu tarihlendirmeyi onaylamakta ve bu türlerin 80 ile 116 milyon yıl arasında genetik ıraksama ile birbirinden ayrıldığını öngörmektedir. Yine bu araştırmaya göre, günümüz köpeksi maymunlara (asıl köpeksi maymunlar ile babun ve şebeklerin dahil olduğu grup) götüren soy ile insansılara götüren soy günümüzden 23 milyon yıl önce, Miyosen başlarında birbirinden ayrılmaya başlamıştır. İnsansılar ise bu yeni tarihlendirmeye göre yaklaşık 15 milyon yıl önce Gibongiller ve büyük insansı maymunlar olmak üzere iki kola ayrılmıştır. Büyük insansı maymunların Asya'daki türleri (orangutanın atası) ile Afrika'daki türleri arasındaki ayrılma günümüzden 11 milyon yıl önce, goril ile şempanzelerin birbirinden ayrılması 6,5 milyon yıl önce ve son olarak şempanze ve hominini ayrılması ise 5,2 (± 1,1) milyon yıl öncesine denk gelmektedir.
Bu zamanların şu anda ne kadar geçerli olduklarını aşağıdaki örnekler göstermektedir: Moleküler saat yöntemi ile yapılan hesaplamalarda primatların ortaya çıkışı 90 milyon yıl önceki Kretase dönemine tarihlendirilirken bulunan fosil kayıtlar temelinde yapılan hesaplamada ise 56 milyon yıl öncesine, Paleosen dönemine tarihlendirilmiştir. Daha önceki araştırmalarda temel olarak alınan mutasyon oranları, goril genom diziliminin çözülmesinden sonra goril ile şempanzenin ayrışma zamanını 5,95 milyon yıl olarak gösterirken fosil bulgular temelinde yapılan çalışma ise bu tarihlendirmeyi göreceli hale getirmiş ve -günümüzde yaşayan Homo sapiens popülasyonlarının mutasyon oranları temel alınarak- soy çizgilerinin ayrımını 6-10 milyon yıl önce gerçekleştiğini gösteren daha yavaş bir mutasyon oranı öngörmüştür. 2010 yılı başlarında Terry Harrison fosil kalıntılar yardımıyla şempanze ve hominini ayrımını 7,5 milyon yıl öncesine tarihlendirirkenClaude Owen Lovejoy ise 2009 yılında, moleküler saat temelinde yapılan çalışmasında şempanze insan ayrımını 6 ile 5 milyon yıl öncesine tarihlendirmiştir.
İnsan ve diğer primatların kromozom yapılarıyla ilgili genom projelerinin tamamlanması, bu türlerin ortak ata ve DNA özgeçmişlerine işaret eden bulgular sunmuştur. (Görsel kaynak[])
İnsandan önce

Primatların kökeni

Fosil kayıtlarından izlenebilen ve primatların atası olduğu düşünülen plasentalı memelilenin orman tabanında yaşayan yumuşakcalarla beslenen bir ağaç sivri faresi benzeri (örn. Zalambdalestes) canlı olduğu görüşü yaygındır. Bu canlı ~70 milyon yıl önce orman tabanını bırakarak orman tavanına yani ağaçlara sıçradı. Uzmanlarca bu canlıya ağaç sivri faresi denmiştir.
İnsanı oluşturacak ağaç sivri faresinin yaklaşık 60 milyon yıl boyunca ağaçlarda yaşadığı düşünülüyor. Bu süre sonunda bedeni irileşmiş, iskeleti dikilme yönünde gelişmiş, parmakları nesneleri güçlü bir şekilde kavrayacak şekilde gelişmiş, gözleri üç boyutlu olarak görme olanağı verecek şekilde yanlardan önlere doğru kaymıştır. Bilinen en eski muhtemel primat, ABD, Montana'da 66 milyon yıl önce yaşamış Purgatorius'tur.
Bazı bazal primatlar
- Notharctus: 50 milyon yıl önce Kuzey Amerika'da yaşamış lemur benzeri bir adapiform cinsi, lemurların veya tüm nemli burunlu maymunların muhtemel atasıdır.
- Altiatlasius: Fas'ta birkaç azı dişi ve bir çene parçası bulunmuş olan bu cins 57-56 milyon yıl öncesine tarihlenmiş olup, birçok paleontolog tarafından ilk gerçek primat olduğu düşünülüyor.
- Teilhardina: Kuzey Amerika ve Asya'da bulunan fosilleri 56 milyon yıl öncesine tarihlenmiş bir omomyid cinsi.
Primatların erken evrimi
Primatların evrimsel tarihi günümüzden 65 milyon yıl öncesine kadar takip edilebilir. Bilinen en eski primat benzeri memeli türler olan plesiadapiformlar, Kuzey Amerika'dan gelmekle beraber Paleosen ve Eosen devrinde elverişli tropikal iklim şartlarında Afrika ve Avrasya'ya kadar yayılmışlardı.
Modern iklim şartlarının başlangıcı, günümüzden yaklaşık 30 milyon öncesine uzanan Erken Oligosen dönemde ilk antarktik buzulların oluşumu tarafından tetiklenir. Bu döneme ait bir primat örneği Notharctus'tur. 1980'li yıllarda Almanya'da bulunan 16,5 milyon yıllık fosil, Doğu Afrika'daki benzer formlardan 1,5 milyon yıl daha eski olup insanların en eski atalarının Afrika kökenli olduğuna dair teorileri sorgulamaktadır.
Bir görüşe göre, Avrasya coğrafyasında yayılarak gelişen bu primatlar Dryopitekus da dahil olmak üzere Afrikalı büyük insansı maymunlar ile insanlara giden soyu oluşturdular ve bu soy daha sonra Avrupa kıtasından Batı Asya'ya ve Afrika'ya doğru göç ettiler.
İnsansıların evrimsel gelişimi
İnsansıların üst ailesi olan Hominoidea'ların beşiği, günümüzden 23 ile 16 milyon yıl öncesine yayılan. erken Miyosen döneminin Doğu Afrikasıdır. Hominoidea üst ailesinin erken Miyosen dönemdeki ilk formları "arkaik" veya "kök Hominoidea" olarak tanımlanır. Bu ilk formların birbirleriyle olan akrabalık ilişkileri ve bu ilk formlar ile onların daha sonra ortaya çıkan türleri arasındaki akrabalıkları fosil örneklerin henüz yetersiz sayıda bulunmuş olmalarından dolayı tartışmalıdır. Bunun yanında bilinen Miyosen dönemi ilk insansıların hiçbirisi, büyük insansı maymunlar için yürümeye dair tipik olan fiziksel özellikleri henüz göstermemektedirler. Görünüşe göre, Miyosen dönemi insansıları artan oranlarda ormanlardan açık alanlara çıkarak daha sonra günümüzden yaklaşık 5 milyon yıl önce Pliyosen devri insansılarını (Hominoidea) oluşturmuşlardır.

Bundan yaklaşık 18 ile 15 milyon yıl önce iki evrimsel kol ayrılır: Bir kol gibonlara (Hylobatidae) ayrılırken diğer kol halihazırda Miyosen döneminde soyları tükenmiş olan cinslere, Afropitekus, Kenyapitekus, Griphopitekus, Pierolapitekus, Dryopitekus ve Oreopitekus'a ve ayrıca büyük insansı maymunlara ayrılır; yani Asya'daki orangutanlar (Ponginae) ile Afrika'daki büyük insansı maymunlara (Homininae: goril, şempanze, insan).
Bazı uzmanlar paleontolojik bulgulardan yola çıkarak büyük insansı maymunların, bu dönemde var olan ve Proconsul'a ismini veren Proconsulgiller üst ailesinden (Proconsuloidea) gelmiş olduğunu varsayarlar. Nitekim Proconsuloidea ailesi türler açısından son derece zengin ve sıra dışı bir takson oldukları gibi özellikle Afrika'nın tropikal yağmur ormanlarında ve Arap Yarımadası'nda bulunmuşlardır. Diğer paleontologlar ise Proconsulgillerin bir kardeş taksonunun büyük insansı maymunları oluşturmuş olabileceği görüşünü daha olasılıklı bularak buna dair herhangi bir spekülasyonda bulunmazlar. Şimdiki bilgi düzeyine göre, bilinen Miyosen dönemi Hominoidea taksonlarından hiçbiri Afrika büyük insansı maymunları ve Hominini soyu için doğrudan bir ortak ata olarak gösterilememektedir.
Avrasya'daki dağılımları

17 ile 14 milyon yıl önce Afrika kliması Rift Vadisi'nin tektonik yükselişi etkisiyle kurumaya başlarken diğer yandan yıllık sıcaklık ortalamaları arasında güçlü farklılıklar da ortaya çıkmaya başladı. Ekolojik şartlardaki bu değişimler, bugünkü bilgilerimize göre Proconsulgillerde çeşitliliğin azalmasına yol açarken köpeksi maymungiller ile insansılar (örneğin Kenyapithecus wickeri, Equatorius africanus ve Nacholapitekus) bu tarihlerde hakim olan ve çoğunluğa geçen cinsler olmaya başladılar. Bu dönemden kalma fosillerde, güçlü çene yapıları, kalın azı dişleri ve diş minesi kalınlığının artışı gibi sert kabuklu ve lifli yiyeceklere uyum sağlama ile ilgili adaptasyonların geliştirilmiş olduğuna ve 16 ile 15 milyon yıl önce ilk defa Afrika dışında kalan yerlerde, Avrasya'nın geniş bölgelerinde bu türlerin yerleşmiş olduğuna dair izlere rastlamak mümkündür.
Yaklaşık 16 milyon yıl önce, erken Miyosen'den orta Miyosen dönemine girildiğinde orangutanın Asya'daki türleri diğer büyük insansı maymunlardan ayrılmaya başladı. Orangutanlar ile akraba olan gruplar içinde fosil kalıntılardan bildiğimiz Ramapitekus, Sivapitekus, Ankarapitekus, Lufengpitekus, Khoratpitekus ve Gigantopitekus gibi cinsler yer almaktadır. 1930'lu yılların başlarında Kuzey Hindistan'da ilk defa fosil kalıntıları bulunan Ramapitekus, 1960 ve 1970'li yıllarda hatalı bir biçimde Homininlerin atası olarak düşünülmüştü.

13 ile 9 milyon yıl önce insansı türlerin sayısı Avrupa kıtasında artmaya başlamıştır. Bu döneme ait İspanya'da bulunmuş olan Pierolapitekus ve Anoiapitekus dışında en az dört ayrı Dryopitekus türünün daha olduğu da bilinmektedir. Buna benzer bir gelişme, tek hayatta kalanının orangutan olduğu Asya'daki diğer formlarda da görüldü.
9,6 milyon yıl önce Avrupa'da Valesiyen krizi olarak bilinen olayda ekosistemde önemli değişiklikler baş göstermeye başladı. İspanya'nın Akdeniz bölgesinden başlayarak Batı ve Orta Avrupa'nın subtropikal ve daima yeşil ormanları giderek artan soğukluk nedeniyle yok oldular. Bu orman örtüsünün yerini yaprak döken ağaç türleri ve güneydeki bazı bölgelerde de bozkırlar ve stepler aldı. Bitki örtüsünü etkileyen bu iklim değişiklikleri bölgede yaşayan hayvanların yaşamını da önemli bir şekilde etkilemeye başladı. Bunun sonucu olarak bu dönemdeki çoğu insansıların Avrupa'daki türleri yok oldular. Sadece Oreopitekus, yaklaşık 7 veya 6 milyon yıl öncesine kadar Sardinya / Korsika bölgesindeki bir adada hayatta kalmayı başardı. 8 ile 7 milyon yıl önce Tibet Platosu'nun tektonik yükselişi Asya'da muson iklim yoğunluğunun artmasına neden oldu. Bunun sonucu olarak, C4-bitki türlerin artış göstermesiyle kanıtlandığı üzere -Avrupa da dahil- yağış sıklığı azalmaya başladı. İklim değişikliği nihayetinde Avrupa'daki tüm insansıların neslinin tükenmesine yol açarken Asya'da da sadece orangutanlar ile gibongillerin hayatta kalabildiği şiddetli bir tür azalmasına neden oldu.
Afrika'daki gelişim
Kuzey ve Doğu Afrika da 9,6 milyon yıl önceki Valesiyen krizinden etkilenmiştir. Ancak bu bölgelerde 13 ile 7 milyon yıl öncesine ait fosil kalıntılara pek rastlanılmamaktadır. Bu bölgelerde fosil buluntulara pek sık rastlanılmaması, insansı maymunların Asya'da geliştiği ve daha sonra Afrika'ya geri göç ettikleri var sayımına yol açmıştı. Aslına bakılırsa fosil oluşumu için iklimin çok nemli oluşu gibi elverişsiz şartlar Afrika'da ancak az sayıda fosil bulunmuş olmasının temel nedenlerinden biridir. Ancak 2007'de ilk kez tanımlanan 10 milyon yıllık Nakalipithecus nakayamai'nin Kenya'da bulunması, ayrıca daha önce de Kenya'da olduğu bilinen ve biraz daha eski olan Samburupithecus kiptalami ile Etiyopya'da bulunan ve yine biraz daha eski olan Chororapithecus fosiline rastlanılması ile Afrika'da da Orta ve Geç Miyosen dönemlerinde çok sayıda insansı türlerin yaşamış olduğu ortaya çıkarılabilmiştir. Yaklaşık 6 ile 8 milyon yıl önce Afrika'da Geç Miyosen döneminde goril ve şempanze soy çizgileri Hominini soy çizgisinden ayrılmaya başlamıştır.
Haziran 2006'da Nature dergisinde Broad Enstitüsü, Massachusetts Teknoloji Enstitüsü ve Harvard Üniversitesi tarafından ortaklaşa yayınlanan bir gen analizi, erken Hominini türlerin şimdiye kadar sanılandan daha alışılmadık şekilde bir evrimsel gelişim geçirdikleri göstermiştir. Bu çalışmada 20 milyon insan DNA baz çifti ile şempanze ve goril DNA'ları moleküler saat aracılığıyla birbirleriyle karşılaştırılmış ve çalışma, ilk büyük insansı maymun türlerinden birinin halihazırda 10 milyon yıl önce Hominini'nin ön atalarından ayrıldığı sonucuna varmıştır. Buna rağmen her iki popülasyon birkaç yüz bin yıl sonra tekrar bir araya gelerek karışmış ve melez popülasyonlar oluşturmuştur.
Araştırmacıların ortaya çıkardığı bu sonuca göre, birbirinden ayrılma ve birleşmeler ve yeniden melezleşmeler 4 milyon yıl boyunca yer değiştirerek birbirlerini takip etmiş ve sonunda şempanzelerin ataları 6,3 ile 5,4 milyon yıl önce nihai olarak Hominini oymağından ayrılmıştır. Bu en son gen alışverişi, X Kromozomunun daha sonra insanlar için karakteristik olan bir forma dönmesi ve aynı zamanda yakın bir şekilde şempanze X kromozomunu da andırması, bunun yanında özellikle X kromozomunun kesintisiz olarak çok genç bir yaşa sahip olması nedeniyle kanıtlanmış görünüyor. Ancak bu seneryoya da itirazlar olmuştur.
Hominin türlerinin ayrışması
Hominini grubuna ait cins ve türlerin varoluş süreleri ve hangi zaman aralıklarında yaşadıkları tahmini değerler olup fosillerin bulunduğu katmanların jeolojik analizlerine, yani fosillerin elde edildiği mevkilerde yer alan katmanların yaşlarının doğrudan hesaplanılmasına dayanır.Moleküler saat olarak bilinen yöntem, modern insanın Hominini grubu içinde hayatta kalabilen tek tür olması ve genetik materyalin sadece ona en yakın olan Neandertal insanı ile karşılaştırılabileceği için henüz DNA örnekleri elde edilemeyen diğer Hominini cinsleri için kullanılamamaktadır. Günümüzde goril ve şempanzelerin dahil olduğu büyük insansı maymunlar grubu hayatta kalabilmiş olduğu için moleküler saat bu gruplara uygulanabilmektedir.
Fosil buluntuların ancak sınırlı sayıda olması nedeniyle, günümüzden 7 ile 5 milyon yıl önce yaşamış olan Sahelanthropus, Orrorin ve Ardipithecus cinslerinden hangisinin insanın ön atası olduğu ve bu cinslerin tarihsel bir şekilde tam olarak doğru biçimde atasal soy çizgisinde kronolojik sıraya oturulmuş olduğu belirsizdir. Şempanzelerden ayrılan ve Hominini grubu içinde yer alan bu fosiller göreceli olarak iyi bilinmektedir. Bilindiği kadarıyla şempanze soy çizgisinden ayrılan bu fosillerden en eskisi 7 milyon yaşındaki Sahelanthropus tchadensis olup onu 6 milyon yıl yaşındaki Orrorin tugenensis izlemektedir. Diğerleri ise aşağıda verilmiştir:
- Ardipithecus (5,5 - 4,4 milyon yıl önce) ile onun türleri Ar. kadabba ve Ar. ramidus;
- Australopithecus (4 – 1,8 milyon yıl önce) ile onun türleri Au. anamensis, Au. afarensis, Au. africanus, Au. bahrelghazali, Au. garhi, ve Au. sediba;
- Kenyanthropus (3 – 2,7 milyon yıl önce) ile onun bilinen tek türü Kenyanthropus platyops;
- Paranthropus (3 – 1,2 milyon yıl önce) ile onun türleri P. aethiopicus, P. boisei ve P. robustus;
- Homo (2 milyon yıl önce – günümüz) ile onun türleri Homo habilis, Homo rudolfensis, Homo ergaster, Homo georgicus, Homo antecessor, Homo cepranensis, Homo erectus, Homo heidelbergensis, Homo rhodesiensis, Homo neanderthalensis, Homo sapiens idaltu, Arkaik Homo sapiens, Homo floresiensis.
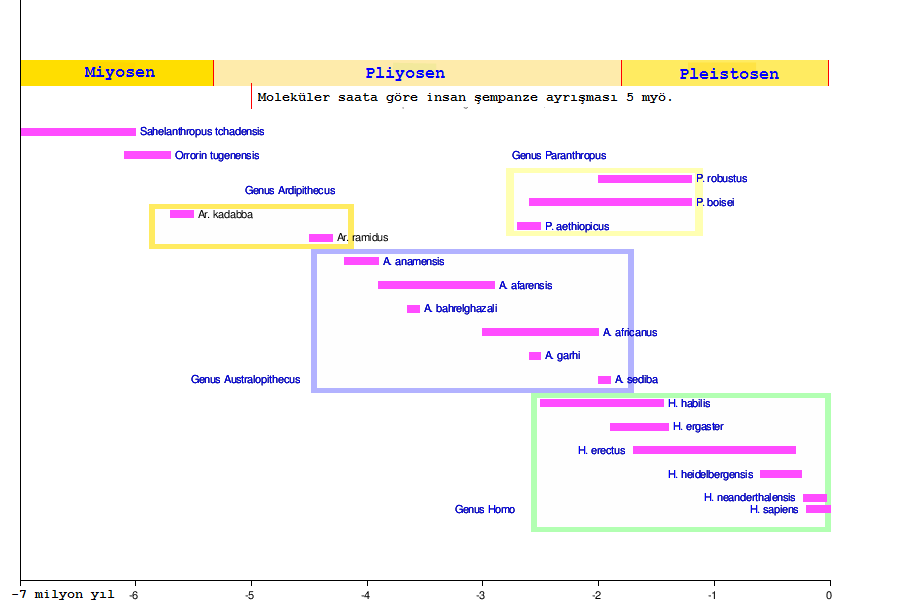
Artan ve azalan oranlarda bulunan fosillerin kronolojik sıralaması onların akrabalık ilişkileri hakkında tam bir vargıya götürmemektedir.

Sahelanthropus tchadensis

Ardipithecus

Ardipithecus

Sahelanthropus tchadensis

Taung çocuğu





Erken dönem Homininleri

Yaklaşık 6 milyon yıl yaşındaki Orrorin tugenensis'in 2000 yılında ve 6 - 7 milyon yıl yaşındaki Sahelanthropus tchadensis fosilinin ise 2001 yılında bulunmalarının ardından her iki türün halihazırda iyi ayak üzerinde yürüyebilen en eski Hominini türleri olduğu açığa çıkmış, böylece insanın doğrudan atası olarak konumlandırıldılar. Ancak bu bulgu, moleküler saatten elde edilen ve Homo ile şempanze soy çizgisinin günümüzden 5 ile 6 milyon yıl önce ayrılmış olduğunu hesaplayan bulguyla örtüşmemektedir.
Daha önce 1994 yılında Etiyopya'da halihazırda Ardipithecus ramidus'e ait fosil kalıntılar bulunmuştu. 4,4 milyon yıl ile tarihlendirilen bu fosiller de birçok araştırmacı tarafından insanın doğrudan atası olarak belirlenmişti. En başta özellikle "Ardi" fosili olmak üzere, bu türün bireyleri de iyi ayak üzerinde yürüyebilme özellikleri göstermektedir. Sahelanthropus, Orrorin ve Ardipithecus'un birbirleriyle olan akrabalık ilişkisi ile daha sonra ortaya çıkan Hominini türleri arasındaki akrabalık dereceleri tartışmalı bir durumdadır ve tam olarak bilinmemektedir.
Bu fosiller, Dünyanın ortalama ısısının günümüzden 4 °C daha sıcak olduğu bir dönemden, Messiniyen ve Zankliyen alt dönemlerindeki geçiş aşamasından gelmektedir. 2001 yılında yayımlanan bir araştırmaya göre, ilk Hominini türlerinin yaşam alanı en çok yüzde 40'ı ağaçlarla kaplı olan, tropik yağmur ormanları ile kuru çöller arasındaki geçiş bölgesinde yer alan geniş çayırlar ve savanlardı. Bu bölgelerde daha sonra, -günümüzden 3,6 milyon yıl önce- ağaç örtüsü biraz daha artarak yüzde 40 ile 60'a ulaşmış; ancak Pliyosen'den Pleistosen'e (Buzul Çağı) geçiş döneminde tekrar azalarak günümüzden 1,9 milyon yıl önce, ağaç örtüsünün yarısından çoğunun, yüzde 50'den daha fazlasının yok olması şeklinde devam etmiştir.
| ERKEN DÖNEM HOMİNİN TÜRLERİ | ||||||
| Görsel | İsim | Yaş | Tür | Bulunuş tarihi |
Ülke | Keşfeden kişiler |
|---|---|---|---|---|---|---|

|
TM 266 (Toumai) | 7 milyon | Sahelanthropus tchadensis | 2001 | Çad | Alain Beauvilain, Fanone Gongdibe, Mahamat Adoum ve Ahounta Djimdoumalbaye |
|
BAR 1000'00 | 6 milyon | Orrorin tugenensis | 2000 | Kenya | Martin Pickford, Kiptalam Cheboi, Dominique Gommery, Pierre Mein, Brigitte Senut |

|
Ardi | 4,4 milyon | Ardipithecus ramidus | 1994 | Etyopya | Yohannes Haile Selassie |
Australopitesinler
Fosil bulgular aracılığıyla hakkında göreli bilgilere sahip olunan ve halihazırda iki ayak üzerinde dik yürüyebilen insanın atalarından biri de Australopitekus'lardır. Australopitekus türlerinin birbirleriyle olan akrabalıkları ve Homo türleriyle olan akrabalık ilişkileri henüz tam olarak açıklık kazanmamıştır. Şu an için Australopithecus anamensis kuşkusuz en erken Hominina türü olarak kabul edilmektedir. Türkçe çevirisi tam anlamıyla "Kuyruksuz Güney Maymun Adamı" olan Australopitekus buluntularına dair en önemli arkeolojik alanlarından biri Güney Afrika'da Johannesburg kentinin 50 km kuzeyinde yer alan Gauteng vilayeti olup zengin fosil yatakları içeren bu bölge "insanlığın beşiği" olarak kabul edilir.UNESCO tarafından dünya mirası kabul edilen insanlığın beşiğinde bugüne kadar gün ışığına çıkarılan fosiller arasında her ikisi Australopithecus africanus türüne ait olan Taung çocuğu ile Dikika çocuğu (Selam) olarak tanınan üç yaşındaki bir kız çocuğu ile Australopithecus afarensis türüne ait olan Lucy fosili bulunmaktadır. Tanzanya'nın Laetoli bölgesindeki Olduvai Kanyonu'nda birkaç Australopithecus afarensis bireyine ait bulunan fosil ayak izleri bu türün temsilcilerinin halihazırda iki ayak üzerinde yürüdüklerini göstermektedir. Buna karşın 1999'da Meave Leakey tarafından Kenya'da bulunan Kenyanthropus platyops'un Australopithecus cinsine mi dahil edileceği yoksa kendi başına ayrı bir cins olarak mı sayılması gerektiği kesinlik kazanmamıştır.
Buna karşın çok güçlü bir çene yapısına sahip olan Paranthropus ise kendi başına bir cins olarak kabul edilmiş ve iri yapılı Australopitesin'lere dahil edilmiştir.Paranthropus gibi iri yapılı Australopitesinlerde güçlü çene oluşumlarına dair adaptasyonların nedeni olarak 2,5 milyon yıl önce ortaya çıkan iklim değişiklikleri ve buna bağlı olarak ısı düşüşleri gösterilmektedir. Bu dönemde kendini gösteren iklim değişiklikleri bir yandan 2,7 milyon yıl önce başlayan arktik Kuvaterner Buzul Çağı, diğer yandan Büyük Rift Vadisi'nin tektonik yükselişi tarafından tetiklendi. Bu değişimlerin sonucu olarak Paranthropus'un atalarının yaşadığı bölgede yağış miktarı azalmaya ve kuraklık dolayısıyla stepleşme olayları görülmeye başladı. İklimin giderek soğumaya ve kurumaya başlamasıyla besin ve yiyecek kıtlığı da başladı. Kısa bir zaman içinde besin olarak ataları taze yapraklarla beslenen bu insansı türler için sadece çok sert liflere sahip olan, katı ve zor sindirilebilir bitkilerden başka bir şey kalmamıştı. Bu kurak bozkırlar, öncelikle kuru çalı veya otlarla beslenebilen ve daha önce küçük formlardaki ataları yaprakla beslenen orman sakinleri olan çift toynaklı ve geviş getiren hayvanlar için besin sunabiliyordu. Bu türler yakın bir zaman içinde büyük sürüler halinde görülmeye başladı ve otçulların sayıları arttığı için etle beslenen veya leş yiyen türler de çoğaldılar. Kendileri için yeterli yiyecek bulamayan dönemin tipik Australopitekusları eğer hayatta kalmak istiyordu iseler bu takdirde yaşamak için çevrelerine uyum sağlayarak değişmek veya yeni beceriler geliştirmek zorundaydılar. Bu şekilde narin yapılı ve iri yapılı Australopitekuslar olmak üzere iki farklı Hominini türü ortaya çıkmaya başladı.
Bazı Australopitekuslar sert lifli bitkileri çiğneyip öğütebilmek için çok kuvvetli çeneler ve güçlü kafatası yapısı geliştirdi. Bunun sonuçlarından biri iri yapılı Australopitekuslara dahil edilen Paranthropus oldu. Diğerleri sorunları çözmek için beyin ve teknik kullandı. Bunlar yiyecekleri daha küçük parçalara ayırmak için ellerine ilk olarak taş aletler aldılar. Yeni yaşama ortamına ayak uyduramayan ve adapte olamayan diğer tüm türler yavaş yavaş yok olmaya başladı.
Paranthropus'un Doğu ve Güney Afrika’da ağaçların seyrek olduğu, sadece kuru ve sert lifli bitkilerin bulunduğu kurak bozkırlarda yaşaması, onun çevresine uyum sağlamasını tetikleyerek onu belirgin bir şekilde narin yapılı diğer Australopithecine türlerinden farklılaşmasına ve diğerlerinden ayrılmasına yol açtı.Paranthropus boisei, Paranthropus robustus ve Paranthropus aethiopicus yaşadığı bu ekolojik niş içinde çok güçlü çiğneme kasları ile çene yapısı ve iri azı dişleri geliştirdi. Bunun yanında kafatasının hemen üstünde yer alan şişkin ve kalın bir kemik tepe çıkıntısı onun çiğneme kaslarını desteklerken güçlü dişleri ile sert kabuklu besinleri kırmasında da önemli rol oynuyordu.
Paranthropus gibi sert lifli bitkileri çiğneyip öğütebilmek için güçlü çene ve diş yapısı geliştirememiş olan diğer narin yapılı Australopitekus türleri ise bunun yerine gitgide daha çok etle beslenmeye yöneldiler. Narin yapılı Australopitekus türleri bu dönemde henüz avcı olarak daha büyük hayvanları avlayamadıkları, bunun yanında avlanmak için sivri pençe veya dişlere de sahip olmadıkları için bu dönemde daha ziyade geride kalan av hayvanlarının artıklarıyla veya taze leşlerle beslenebiliyorlardı. Büyük olasılıkla kemik içindeki iliklere ulaşmak veya sert kabuklu yiyecekleri kırmak için ilk kez bu dönemde taş alet kullanmaya başladılar.
Bunun yanında hangi türün veya cinsin ilk olarak taş alet kullanmış olduğu henüz açıklığa kavuşmamıştır. Bazı araştırmacılar Oldovan taş aletlerinin ilk kez Australopitekuslar tarafından yapıldığını, bunun yanında taş alet yapımının sadece insansı türlere özgü olan bir beceri olamayacağını öne sürerler. Buna göre diğer Australopithecine türlerinin de kaba şekilde olsa dahi taş işlemeye yatkın olmuş olmaları kuvvetle muhtemeldir. Bu varsayımlara dair henüz herhangi bir bulgu olmamakla birlikte ilk Homo türlerinin diğer aletler yardımıyla sistematik biçimde ve geniş çapta farklı taş aletler oluşturabildiklerine dair bulgular mevcuttur.
| NARİN YAPILI AUSTRALOPİTEKUSLAR | ||||
| Tür ismi | Yaşadığı zaman | Yaşadığı yer | Açıklama | Görsel |
|---|---|---|---|---|
| Australopithecus anamensis | 4,2 - 3,9 milyon yıl önce |
Doğu Afrika
|
Dişleri ve çene yapısı ilkel maymun fosillerine oldukça benzerlik göstermektedir. Bulunan bir kaval kemiği parçası onun iki ayak üzerinde durabildiğinin güçlü bir kanıtıdır. Üst kol kemiği ise insanınkine oldukça benzemektedir. | |
| Australopithecus afarensis | 3,6 - 2,9 milyon yıl önce | Doğu Afrika | Düşük alnı, gözün üzerindeki kemik çizgisi, düz burnu ve çenesiz yüzü ile maymuna benzemektedir. Cranial kapasitesi 375-550 cl arasındadır. Çene yapısı maymunların dikdörtgen çenesi ile insanın parabolik çenesi arasında bir form gösterir. Buna bağlı olarak insana benzer dişlere sahip olan afarensisin iskelet yapısı şempanzeyi andırmaktadır. Ancak, leğen ve bacak kemiklerinin insana benzer özellikleri onun iki ayaklı olduğunu kesinleştirmektedir. | |
| Kenyanthropus platyops | 3,5 - 3,2 milyon yıl önce | Doğu Afrika | Bulunan fosil kalıntılarda muhtemelen dik yürüdüğünü düşündürecek ayak kemiği yapısı ile geniş bir düz yüze sahiptir. Dişi tipik insan ve tipik maymun arasında ara geçiş formu gösterir. Kenyanthropus platyops ismi "Kenyalı düz yüzlü adam" anlamına gelir. | |
| Australopithecus africanus | 3 - 2 milyon yıl önce |
Güney Afrika
|
İki ayak üzerinde yürüyebilen, ön kolları bacaklardan biraz daha büyük olan insansı bir tür. Beyin hacmi 420-500 cl arasında olup A. afarensis'ten biraz daha büyüktür. Beyin hacmi şempanzenin beyninden daha büyük olsa da henüz konuşabilecek kadar gelişmemiştir. A. africanus'un diş ve çene yapısı maymunlardan çok insana yakınlık gösterir. Çene yapısı artık tam bir parabol şeklindedir. Azı dişleri A. afarensis'ten daha büyüktür. | |
| Australopithecus garhi | 2,5 milyon yıl önce |
Doğu Afrika
|
Bulunan fosil kafatası (cranium), diğer dört iskelet parçası bu türün diğer Australopithecus türlerine kıyasla daha uzun kalça kemiklere sahip olduğunu göstermektedir. Uzun ve güçlü kollara dair yapıların bulunuşu, bipedal yürümede uzun adımlar atabilmeye doğru giden bir eğilim olduğunu gösteriyor. Büyük azı dişleri Australopithecus aethiopicus'a, yüz, kafatası ve dişler ise Paranthropus'a benzememektedir. | |
| İRİ YAPILI AUSTRALOPİTEKUSLAR | ||||
| Tür ismi | Yaşadığı zaman | Yaşadığı yer | Açıklama | Görsel |
| Paranthropus aethiopicus | 2,7 - 2,3 milyon yıl önce | Doğu Afrika | İlkel ve gelişmiş özellikleri bir arada taşıyan P. aethiopicus'un beyin hacmi 410 cl olup diğer türlere göre oldukça küçüktür ve iskeletinin ilksel özellikler taşıyan bazı kısımları A. afarensis'e benzemektedir. Ancak yüzünün iriliği ve bilinen insansılar içerisinde en büyük kemik tepe çıkıntısına sahip olması onu diğerlerinden ayırır. | |
| Paranthropus robustus | 2,0 - 1,2 milyon yıl önce |
Güney Afrika
|
Yüzü oldukça iri ve düz bir yapıya sahip olup alın yok denecek kadar yatık ve geniş kas çizgilerine sahiptir. Besinleri genelde sert ve çok çiğnemeyi gerektirecek şekilde olduğundan görece küçük ön dişleri ve iri azı dişleri bulunmaktadır. Ortalama beyin kapasitesi 530 cl civarında olup P. robustus iskeletlerinin yanında bulunan kemikler, onların bu kemikleri kazıcı alet olarak kullandığını işaret etmektedir. | |
| Paranthropus boisei | 2,3 - 1,4 milyon yıl önce |
Doğu Afrika
|
P. robustus'a benzemekle beraber yüzü, yan ve arka azı dişleri daha iri yapılıdır. Beyin hacmi yaklaşık 530cl civarında olup erkek bireyler ortalama 68 kg ağırlığında ve 1,3 m uzunluğundayken dişi bireyler 45 kg ve 1,05 m uzunluğundadır. P. robustus tüm Australopitekus türleri arasında en büyük diş ile en kalın diş minesine sahip olan tür olup "fındık kıran" adıyla da bilinir. | |







İlk Homo türlerinin ortaya çıkışı

Yaklaşık 3 ile 2 milyon yıl önce Australopithecus'un bir türünden, özellikle alet kullanımına dair bulgulara rastlanıldığı için fosil kalıntıları Homo cinsine dahil edilen insan soyunun ilk temsilcileri evrildi. Homo cinsinin günümüzde hayatta kalan tek türü Homo sapiens olup nesli tükenmiş olan Homo türlerinden bazıları insanın atası, bazıları ise daha çok modern insanın farklılaşarak atasal soydan ayrılan kuzenleri olarak görülmektedir. Bu gruplardan hangilerinin ayrı bir tür, hangilerinin ise alt tür olarak sayılması gerektiğine dair henüz tam olarak bir görüş birliği sağlanamamıştır. Bunun nedeni bazı durumlarda fosil sayısının az oluşu, bazı durumlarda ise Homo türlerini sınıflandırılmasına dair yaklaşımlar arasında küçük farklılıklar olmasından kaynaklanmaktadır.
Bir Australopithecus türünden ilk Homo türlerinin evrimi, daha önce göreli nemli ve göreli kuraklık arasında sık sık değişiklik gösteren Doğu Afrika ikliminin son bir kez daha kurak iklime doğru değiştiği bir dönemde gerçekleşmiş olduğu için dikkat çekicidir. Bu durum günümüzden 2,8 milyon yıl öncesi ile en az 1,8 milyon en çoksa 1,6 milyon yıl öncesine dayanan sedimentlerdeki toz birikimleriyle, bunun yanında savanların genişlemesi ve Antilop gibi Boynuzlugillere ait çok sayıda hayvan fosillerinin bulunmuş olmasıyla belgelenmiş olup tortul tabakadaki olası en büyük toz çökeltileri ile Homo erectus'un ortaya çıkması aynı döneme rastlamaktadır. Bunun yanında arkeolojik ve paleontolojik bulgulara dayanarak, ilk insanların beslenme alışkanlıkları ve bu beslenme alışkanlıklarının insanın fiziksel evrimi ve davranış farklılıkları üzerinde oynadığı rolün incelenmesiyle ilk Homo türleri arasındaki bu varyasyon farklılıkların açıklanabilmesi bir ölçüde mümkün olmuştur.
Homo rudolfensis ve Homo habilis

Homo cinsinin en eski iki temsilcisi, bugün Kenya'da Turkana Gölü olarak bilinen eski adıyla Rudolf Gölü'nün ismini taşıyan Homo rudolfensis ile Homo habilis'tir. Günümüzden 2,4 ile 1,4 milyon yıl önce yaşamış olan Homo habilis Güney ve Doğu Afrika'da Pliyosen ve Pleistosen dönemlerinde 2,5 ile 2 milyon yıl önce Australopithecine'lerden ayrılarak evrilmiştir. Homo rudolfensis ise günümüzden 1,9 ile 1,6 milyon yıl önce Kenya'da Turkana Gölü çevresinde yaşamıştı.
Bu iki türün birbirleriyle olan akrabalık ilişkisi ile kendinden önce ve kendinden sonra gelen Hominini türleriyle olan akrabalıkları henüz tartışmalıdır. Bu iki türün Australopithecus'a olan anatomik yakınlıkları Bernard Wood'u 1999 yılında onları Australopithecus rudolfensis ve Australopithecus habilis olarak tekrar adlandırılması önerisine yönlendirmiştir. Buna rağmen her iki türün anatomik özellikleri onları genel anlamda daha eski olan Australopithecus türlerinden ayırmak için tanımlanır.
Bu anlamda Homo habilis ile Homo rudolfensis'in yüzleri Australopitekus'ta olduğundan daha basit yapılı olmakla beraber Homo habilis'in gözleri örneğin Homo erectus gibi daha sonra gelen diğer Homo türlerine kıyasla birbirinden daha uzak bir mesafede yer alırlar. Buna karşın alın kemiğinde bir uçtan diğer uca doğru kesintisiz bir şekilde uzanan kaş çıkıntısı Homo erectus'ta olduğundan daha az belirgindir. Homo habilis ve Homo rudolfensis'in kafatasları Australopithecus oder Paranthropus'ta olduğu gibi göz çukurlarının hemen arkasından başlayarak daralıp küçülmezler ve bu yüzden daha çok beyin hacmi oluşur.
|
−10 —
–
−9.5 —
–
−9 —
–
−8.5 —
–
−8 —
–
−7.5 —
–
−7 —
–
−6.5 —
–
−6 —
–
−5.5 —
–
−5 —
–
−4.5 —
–
−4 —
–
−3.5 —
–
−3 —
–
−2.5 —
–
−2 —
–
−1.5 —
–
−1 —
–
−0.5 —
–
0 —
|
|
|
||||||||||||||||||
Bunun yanında her iki türün alt ve üst çeneleri Australopithecus'tan daha küçük olup bu nedenle çiğneme kaslarının kafatasıyla birleştiği üst çene bölgesi daha az belirgindir.Friedemann Schrenk'e göre Homo habilis ile Homo rudolfensis'i birbirinden ayıran özellikler, Homo rudolfensis'te daha büyük bir beyin, Homo rudolfensis'in üst çenesinde yer alan küçük azıdişlerde (premolar) 3 adet, Homo habilis'te ise 2 adet diş kökünün bulunması, alt çenede ise Homo rudolfensis'te 2 kök, Homo habilis'te sadece tek 1 kökün olması, Homo rudolfensis'te yirmi yaş dişlerinin Australopitekus'a kıyasla daha küçük olması ama Homo habilis'te küçük olmaması, Homo rudolfensis'in üst bacak kemiği ile ayak yapısı insanlarınkine benzerken Homo habilis'te Australopitekus'a benzemesidir.
Homo gautengensis
Australopitekuslardan daha küçük dişe ve daha büyük beyne sahip olan Homo habilis'in taş ve kemik aletler yapabildiği bilinmekte olup onu keşfeden Louis Leakey tarafından ona "becerikli insan" anlamına gelen Homo habilis ismi konulmuştu. Mayıs 2010 tarihine kadar Homo habilis'in en eski Homo türü olduğu düşünülüyordu; ancak bu tarihte Güney Afrika'da Homo gautengensis'e ait fosil kalıntıların bulunmasıyla bu yeni türün Homo habilis'ten de eski bir tür olduğu anlaşıldı.
İlk defa 1977 yılında Güney Afrika'da Johannesburg yakınlarındaki Sterkfontein mağarasında bulunmuş ancak bugüne kadar ihmal edilerek incelenmemiş olan bu fosil kalıntılar Mayıs 2010'da tekrar Dr. Darren Curnoe tarafından incelenmiş ve bu fosil parçalar arasında Swartkrans birey 1 fosilinin 1,9 ile 1,8 milyon yıl ile en eski olanı, diğer StW 53 isimli fosilin ise 1,8 ile 1,5 milyon yıl olduklarını saptamıştır. İnceleme sonucu Homo gautengensis'in dişlerinin daha büyük olduğu beyninin ise Homo erectus, Homo sapiens, hatta Homo habilis'ten dahi daha küçük olduğu ortaya çıkmıştır. Homo gautengensis'in boyu 0,91 m olup ağırlığı ise 50 kg kadardı. İki ayak üzerinde yürümesine rağmen avcılardan korunmak, uyumak veya beslenmek için ağaçlarda da zaman geçiriyordu.
Homo ergaster ve Homo erectus

Günümüzden yaklaşık 2 milyon yıl önce Homo ergaster ortaya çıktı. Günümüzden 1,5 ile 1 milyon yıl önce, erken Pleistosen dönemde Homo ergaster'in Afrika, Asya ve Avrupa'daki bazı popülasyonları büyük bir beyin ve daha gelişmiş taş aletler geliştirdi. Bu ve diğer farklılıklar Homo ergaster'i diğerlerinden ayrı ve yeni bir tür olarak sınıflandırmak için çoğu paleoantropolog tarafından yeterli görünmektedir. Buna rağmen Homo ergaster'in ayrı bir tür olarak değerlendirmesi hala tartışılmalı olup bazı araştırmacılar Homo ergaster fosillerini Homo erectus türünün ilk örneklerinden biri olarak değerlendirirler. Bu anlamda Homo ergaster'in erken dönemlerde 1,8 ile 1,25 milyon yıl önce, Homo erectus'tan ayrı bir tür olarak ayrılmaya veya Homo erectus ergaster olarak alt türe dönüşmeye başladığı düşünülmektedir.
Homo erectus ise günümüzden 1,8 milyon ile 70.000 yıl önce Afrika'da, Hindistan, Çin ve Cava adası gibi çeşitli bölgelerde yaşamış olup ilk fosil kalıntısı 1891 yılında Eugene Dubois tarafından Endonezya'da Cava adasında bulunmuştu. Homo erectus'un Toba felaketi sonucu yok olduğu düşünülmektedir.Homo erectus, Afrika'dan ilk çıkan Ön Asya üzerinden Doğu Asya'ya ve Avrupa'ya yayılan ilk Homo türüydü. Bununla beraber Homo erectus soloensis ve Homo floresiensis bu felaketten kurtulmuş görünüyor. Bunun yanında Homo erectus gerçek anlamda iki ayak üzerinde dik yürüyen ilk Homo türüdür. Bu da kafatasının alt kısmında bulunan boyun deliği Foramen Magnum'un farklı bir şekilde yerleşmesi ve dizlerinin kilitli bir şekilde evrilmesi sonucu ortaya çıkmıştır. Homo erectus'un ateş yakıp kullanabildiği düşünülmektedir.
En bilinen Homo erectus örneği Pekin Adamı olup diğer örnekleri Asya'da (özellikle Endonezya'da), Afrika ve Avrupa'da bulunmuştur. Bazı paleoantropologlar Asyalı olmayan Homo erectus grupları için Homo ergaster terimini kullanırlar veya Asya'da bulunan ama Homo erectus'tan az farkla ayrılan fosiller için kullanmayı tercih ederler.
Homo georgicus
Homo cinsinin Afrika kıtası dışında bulunan ilk kanıtları Gürcistan'da 1,8 milyon yıl öncesi ile tarihlendirilen ve daha önce Homo georgicus olarak adlandırılan Dmanisi fosil kalıntılarıdır. Dmanisi kazı alanında bulunan ve yaklaşık 1,85 milyon yıl öncesine ait olan 100′ün üstündeki bu taş aletler insanın Asya’daki ön ataları ve buradaki yerleşimleri hakkında en eski ve belgelenmiş örnekleri teşkil etmektedir. Bunun yanında Dmanisi’de bugüne kadar üç kafatası, büyük ölçüde aşınmış azı dişleri, bir de alt çene olmak üzere yirminin üzerinde insan kalıntısı bulunmuştur. Bu bulgulara göre 1,50 m boyundaki Dmanisi insanları, Afrikalı çağdaşlarına göre daha küçük ve narin yapılı olup beyin hacimleri de 0,6 - 0,8 litre arasındaydı. Bu bulgular Homo erectus'un 1,85 milyon yıl önce veya bundan daha erken Afrika’dan çıktığını ve dünyaya yayıldığını gösteriyor.Homo georgicus olarak adlandırılan bu kalıntıların Homo soyağacı içindeki yeri son yapılan araştırmalarla belirlenmemiş olup Dmanisi'de bulunan bu fosiller Homo erectus olarak tanımlanmıştır. Bulunan Homo erectus georgicus fosilleri erkek bireylerin belirgin bir şekilde kadın bireylerden çok daha uzun boylu olduğu için seksüel dimorfizm göstermektedirler. Dmanisi'de bulunan bu kalıntılar ayrı bir Homo türü olarak değil, aksine Homo habilis'in Homo erectus'a dönüşmesinin hemen ardından, 1,8 milyon yıl önce gerçekleşen bir ara form aşaması olarak değerlendirilmektir.
Homo antecessor ve Homo cepranensis
İspanya'da bulunan ve 1,2 milyon yıl öncesi ile tarihlendirilen Homo antecessor fosillerin ayrı bir Homo türü mü olduğu yoksa Homo erectus'un başka bir yerel varyantı mı olduğu açıklık kazanmamıştır. Bazı araştırmacılara göre Homo antecessor, Homo ergaster ile Homo heidelbergensis arasında bir ara geçiş formu olabileceği gibi Richard Klein gibi bazı araştırmacılara göre Homo ergaster'den farklılaşarak ayrılan ayrı bir tür de olabilir.
Bunun gibi İtalya'da 1994 yılında Roma yakınlarında Frosinone ilinde bulunan ve yaklaşık 350.000 ile 500.000 yıl öncesi bir döneme ait olduğu saptanan Homo cepranensis'e ait kafatası kemik kalıntısı da H. erectus ile H. heidelbergensis arasında ara geçiş özellikleri göstermektedir.
Homo heidelbergensis
Günümüzden yaklaşık 800.000 yıl önce Homo erectus'tan, ondan daha büyük bir beyine sahip olan Homo heidelbergensis evrildi.Homo heidelbergensis'e ait en iyi korunmuş fosil kalıntılar 600.000 ile 400.000 yılları öncesine ait olup Heidelbergensis insanına ait taş alet kültürü, Paleolitik Çağ’da Homo sapiens ve Homo erectus'un el baltaları ve yongalardan hazırladıkları ve kesici alet kullanımıyla standartlaştırdıkları Acheuleen kültürüne çok benzemektedir. Homo heidelbergensis ortalama olarak 1,8 m boyunda ve 100 kg ağırlığında olup diğer Homo türlerinde kıyasla biraz daha uzun boyunlu idi. Ona ait en büyük fosil 2,13 m uzunluğunda olup Heidergensis adamı 0,5 milyon ile 300.000 yıl önce öncelikli olarak Güney Afrika'da ve Avrupa kıtasında yaşamıştı.İspanya Atapuerca'da bulunan Homo heildenbergensis fosilleri onun ölülerini gömen ilk Homo türü olduğunu göstermektedir. Bazı araştırmacılara göre, Homo heidelbergensis onların torunları olan Neandertaller gibi ilkel bir dil geliştirmiştir.
Homo heidelbergensis'in Anglo-Amerikan paleontolojisinde yaygın olduğu gibi ayrı bir tür olarak mı yoksa Avrupa paleontolojisindeki gibi Homo erectus'un bir alt türü olarak mı sınıflandırılması gerektiği tartışmalıdır. Afrika'da kalan Homo erectus popülasyonlarından günümüz modern insanı Homo sapiens evrilirken Avrupa'da Homo heidelbergensis'den veya alt tür olarak kabul edilirse Homo erectus heidelbergensis'ten Neandertaller (Homo neanderthalensis) evrilmiştir.
Homo rhodesiensis ve Gawis kafatası
300.000 ile 125.000 yıl öncesine tarihlendirilen Homo rhodesiensis veya Rodezyalı Adam, en güncel araştırmalara göre Homo heidelbergensis grubu içine dahil edilmekle beraber daha önce Arkaik Homo sapiens veya Homo sapiens rhodesiensis olarak da sınıflanırılmıştır. Şubat 2006 tarihinde Etiyopya'nın Orta Afar bölgesinde Gawis yakınlarında bulunan ve bu nedenle Gawis kafatası olarak adlandırılan bir fosil Homo erectus ile Homo sapiens arasında ara özellikler göstermekte olup bu fosilin birçok evrimsel çıkmaz gibi geride bir soy bırakmadığı da düşünülmektedir. Bu kalıntılar 500.000 ile 250.000 yılları arasında tarihlendirilmekte olup fosili keşfedenler tarafından henüz kapsamlı bir araştırma yayınlanmadığı için bu kalıntılar hakkında sadece özetlenmiş bilgiler bulunmaktadır. Gawis adamının yüz özellikleri onun bir ara form ya da dişi bir "Bodo insanı" olduğunu düşündürmektedir.
Neandertal ve Denisova insanı

Homo neanderthalensis alternatif olarak Homo sapiens neanderthalensis olarak da isimlendirilmektedir ve günümüzden 400.000 ile 30.000 yıl önce Avrupa ve Asya'da yaşamıştır. Mitokondriyal DNA dizilemesinden elde edilen kanıtlar Homo neanderthalensis ve Homo sapiens arasında önemli veya dikkate değer bir gen akışı olduğunu göstermediği için bu iki tür 600.000 yıl önce ortak bir ataya sahip olan iki ayrı tür olarak değerlendirilmektedir. Ancak, 2010 yılında yapılan Neandertal insanına ait genom dizilemesi Neandertaller ile anatomik modern insanın gerçekten de 45.000 ile 80.000 yıl önce (modern insanın Afrika'dan çıkıp göç ettiği ama henüz Avrupa, Asya veya diğer bölgelere yayılmamış olduğu dönem) birbirleriyle gen alışverişinde bulunduklarını göstermiştir. Hemen hemen Afrika kıtası dışındaki tüm modern insanlar %1 ile %4 arasında Neandertal geni taşımaktadır ve bu sonuç insanlardaki bazı alellerin ıraksamasını sorgulanmakta olmasına rağmen 1 milyon yıl ile tarihlendiren güncel araştırmalarla da uyumludur.Homo sapiens ile olan rekabet Neandertallerin yok olmasına muhtemelen bir rol oynamıştır.Homo sapiens ve Neandertal insanı 10.000 yıl boyunca Avrupa'da bir arada var olmuş olabilirler. Daha yeni tarihli bir araştırma ise Neandertallerin modern insanlar kadar fazla genetik çeşitliliğe sahip olmadığını, bu nedenle 50 bin yıl önce yok olma tehlikesi yaşadığı, yeni koloniler kurarak 10 bin yıl daha hayatta kalabildiğini, bu anlamda Neandertallerin modern insanın Avrupa'ya ulaşmadan önce halihazırda yok olmaya başladıklarını göstermektedir.
Neandertal insanının yanı sıra günümüzden 40.000 yıl önce Rusya'nın Altay Dağları'nda yer alan Denisova Mağrası'nda Neandertallerin akrabası olan ve Denisova insanı olarak bilinen yeni bir insansı tür ortaya çıktı. Bu insanlardan kalma şimdiye kadar bir azı dişi ile birer el ve ayak parmak kemikleri bulunmuş olup bu kalıntılar bilimsel olarak incelenerek tanımlanmıştır. Bu kalıntılardan elde edilen mtDNA ile nükleit-DNA'larının analizleri sonucu Denisova insanların Hominini grubu içinde en çok Neandertaller ile yakın akraba oldukları, bu verilerin onları yeni bir insansı tür olarak tanımlamak için yeterli olduğu ve Neandertallerin yanında en az 250.000 yıl süren bağımsız bir popülasyon öz geçmişine sahip olduklarını ortaya çıkarmıştır. Yine bu araştırmalarda bazı Melanezya yerlilerinin Denisova insanı ile %6 ortak genlere sahip olduğunu ve Güneydoğu Asya'da sınırlı bir gen alışverişi olduğunu da göstermektedir.
Buna göre yaklaşık 40.000 yıl önce Altay Dağları'nda Homo sapiens ve Neandertaldan başka oraya göç eden üçüncü bir insansı tür daha yaşıyordu. Bunun yanında aynı zaman diliminde, Endonezya'da Flores adasında saptanan ve cüce insan diye bilinen Homo floresiensis türü de yaşamıştı. Cava Adası gibi daha uzak ve tenha bir bölgede 50.000 yıl öncesine ait Homo erectus kalıntılarının bulunmuş olduğu da göz önüne alınırsa Homo cinsine dahil olan bu dört farklı insan türü, -Homo erectus, Neandertaller, Homo floresiensis, modern insan ve Denisova insanı belirli bir zaman dilimi içinde aynı anlarda Avrasya'ya yayılmışlardır.
Homo floresiensis
Popüler ismiyle "hobit" olarak tanınan Homo floresiensis'in 2003 yılında Endonezya'nın Flores Adası'nda bulunan kalıntıları bugün Homo erectus'un daha sonra ada cüceleşmesi yoluyla küçülmüş olan bir formu olarak da yorumlanmaktadır. Bu türün temsilcileri 100.000 ile 12.000 yıl öncesine kadar Endonezya'nın Flores adasında yaşıyorlardı. Homo floresiensis modern insanla ortak olmayan ama modern insandan türeyen özelliklere sahip Homo türünün yeni bir örneği olarak sahip olduğu küçük boyutu ve yaşı ile de ilgi çekmektedir. Başka bir deyimle, Homo floresiensis modern insanla ortak bir ataya sahip olmakla birlikte modern insanın soy çizgisinden ayrılarak bağımsız bir evrimsel yol takip etti. Homo floresiensis'e dair bulunan başlıca kalıntı 30 yaşındaki bir kadına ait iskelet fosilidir. 2003 yılında bulunan fosil 18.000 yıl öncesi ile tarihlendirilmektedir. Flores adasında yaşamış olan kadının boyu 1 metre uzunluğunda, beyin hacmi ise 380 cm3 olup bu değer şempanze beyninden veya ortalama bir Homo sapiens beyninden (1400 cm3) daha küçüktür.
Bununla birlikte Homo floresiensis'in ayrı ve bağımsız bir tür olup olmadığı güncel olarak tartışılmaktadır. Bazı araştırmacılar Homo floresiensis'in cücelik sendromuna sahip bir modern Homo sapiens olduğu görüşündedir. Bu hipotez, Flores adasında yaşayan bazı modern insanların pigme kökenli oldukları için kısmen desteklenmiştir. Bu durum patolojik cücelik ile birleştiğinde cüce-benzeri bir insan oluşturmuş olabilir. Bu hipotezi başlıca destekleyen diğer bir bulgu da Homo floresiensis ile birlikte sadece Homo sapiens ile bağdaşlaştırılabilecek aletlerin bulunmuş olması.
Ancak patolojik cüceliğe dair bu hipotez Homo floresiens'in, hasta olsun olmasın modern insana benzemeyen ama Homo cinsinin daha eski üyelerine benzeyen fazladan ve ilksel özelliklerini açıklamakta yetersiz kalmaktadır. Kafatası özelliklerinin dışında el bileğindeki kemiklerin formu, dirsekle bilek arasında kalan kolun ön kısmı, omuz kemikleri, bunun yanında diz ve ayak kemiklerinin şekli bu hipotezin açıklamakta yeterli olmadığı özelliklerdir.
Homo sapiens
Homo sapiens Latince'de "akıllı, bilge" anlamına gelir ve yaklaşık 250.000 yıl öncesinden günümüze kadar yaşamaktadır. Orta Pleistosen döneminde, yani günümüzden 250.000 yıl önce, 400.000 yıl öncesi ile ikinci Buzul Çağı arasında bir dönemde, Homo erectus'tan Homo sapiens'e doğru geçiş ile birlikte insanın beyin hacminde ve taş alet teknolojisinin gelişiminde büyük bir patlama yaşanmıştır. Şimdiye kadar elde edilen kanıtlar Homo erectus'un Afrika'dan çıkarak diğer bölgelere göç ettiğini, bunun yanında Afrika'da Homo erectus'tan Homo sapiens'e doğru bir türleşme olayı yaşandığını göstermektedir. Afrika içinde ve Afrika kıtası dışına sonradan meydana gelen ikinci bir göç dalgası ile daha önce yayılmış olan Homo erectus'un yerine yenileri gelmiş ve Homo erectus'un gerilemesine ve giderek azalmasına neden olmuştur. İnsan türünün tarih öncesi bu göçleri ve sahip olduğu ilk kökenler genel olarak "tek orijin" veya "Afrika'dan çıkış" teorileri ile ele alınmakta ve ifade edilmektedir. Bunun yanında güncel bulgular insan evriminde aynı zamanda "çoklu bölge" modellerine de imkân tanımakta ve sonradan göç eden Homo sapiens popülasyonlarının diğer yerel Homo türleriyle karışmış olabileceğini de hesaba katmaktadır. Bu güncel olarak paleoantropolojide hararetli tartışmaların sürmesine neden olan bir konudur.
Güncel araştırmalar Homo sapiens'in genetik olarak oldukça homojen bir yapıya sahip olduğunu, bunun yanında bireylerin sahip olduğu DNA'ların diğer canlı türlerinde olduğundan birbirlerine daha çok benzediği, bunun insanın göreceli olarak henüz yeni evrilen bir tür olduğundan veya Toba felaketi gibi doğal felaketler sonucu oluşan olası bir genetik darboğazdan kaynaklanabileceğini göstermektedir. Belirgin genetik özellikler, ilk olarak az sayıda kişilerden oluşan insan gruplarının başka koşullara sahip olan yeni bir çevreye göç etmesi sonucu ortaya çıkmıştır. Çevreye uyum sağlama sonucu ortaya çıkan bu özellikler Homo sapiens genomunun ancak çok küçük bir parçası olup bu çeşitli özellikler arasında deri rengi, burun yapısı veya şekli gibi yüzeysel dış özelliklerin yanında deniz seviyesinden daha yüksek olan rakımlarda daha verimli nefes alabilme yeteneği gibi ilk bakışta gözle görünemeyen iç özellikleri de barındırır.
| HOMO CİNSİNE BAĞLI TÜRLERİN KARŞILAŞTIRILMASI | |||||||
| Tür | Yaşadığı zaman (Milyon yıl önce) | Yaşadığı yer | Yetişkinlerin boyu | Yetişkinlerin ağırlığı | Beyin Hacmi (cm3) | Fosil kayıtları | Cins isminin keşfediliş/kayıtlara geçiş yılları |
|---|---|---|---|---|---|---|---|
| H. habilis | 2,2 – 1,6 | Afrika | 1,0–1,5 m | 33–55 kg | 660 | çok | 1960/1964 |
| H. erectus | 2 – 0,03 | Afrika, Avrasya (Cava Adası, Çin, Kafkaslar) | 1,8 m | 60 kg | 850 (önceleri) – 1100 (sonraki zamanlarda) | çok | 1891/1892 |
| H. rudolfensis | 1,9 | Kenya | 1 adet kafatası | 1972/1986 | |||
| H. georgicus | 1,8 | Gürcistan Cumhuriyeti | 600 | az | 1999/2002 | ||
| H. ergaster | 1,9 – 1,4 | Doğu ve Güney Afrika | 1,9 m | 700–850 | çok | 1975 | |
| H. antecessor | 1,2 – 0,8 | İspanya | 1,75 m | 90 kg | 1000 | 2 bölge | 1997 |
| H. cepranensis | 0,9 – 0,8? | İtalya | 1000 | 1 adet kafatasının üst kısmı | 1994/2003 | ||
| H. heidelbergensis | 0,6 – 0,25 | Avrupa, Afrika, Çin | 1,8 m | 60 kg | 1100–1400 | çok | 1908 |
| H. neanderthalensis | 0,35 – 0,03 | Avrupa, Batı Asya | 1,6 m | 55–70 kg (yapılı bünye) | 1200–1700 | çok | (1829)/1864 |
| H. rhodesiensis | 0,3 – 0,12 | Zambiya | 1300 | oldukça az | 1921 | ||
| H. sapiens sapiens | 0,2 – günümüz | dünya genelinde | 1,4–1,9 m | 50–100 kg | 1000–1850 | halen yaşamakta | —/1758 |
| H. sapiens idaltu | 0,16 – 0,15 | Etiyopya | 1450 | 3 adet kafa kemiği | 1997/2003 | ||
| H. floresiensis | 0,10 – 0,012 | Endonezya | 1,0 m | 25 kg | 400 | 7 birey | 2003/2004 |
İnsanın kökenine dair modeller
Günümüzde yaşayan tüm insanlar, herhangi bir tür bariyeri tarafından bölünmemiş bir şekilde Homo sapiens sapiens'in tek bir popülasyonuna dahildirler. Ancak "Afrika'dan çıkış" modeline göre, Homo sapiens ilk insan türü değildir: Homo cinsine ait ilk tür en az 2 milyon yıl önce Doğu Afrika'da evrilen Homo habilis ve bu türün nispeten kısa bir süre içinde Afrika'nın değişik bölgelerine yayılan üyeleridir. Homo erectus günümüzden 1,8 milyon yıldan biraz daha fazla bir zaman önce evrildi ve 1,5 milyon yıl önce de Avrasya ve Afrika'yı içeren Eski Dünya'nın tüm bölgelerine yayıldı.
Antropologlar, mevcut modern insan nüfusunun sadece Doğu Afrika'da evrildiği ve daha sonra Afrika dışına göç ederek -Afrika'dan çıkış modeli veya tümden yer değiştirme modeli olarak öne sürüldüğü şekilde- Avrasya'daki insan nüfusunu oluşturduğunu veya Çoklu bölge modelinin ileri sürdüğü gibi birbirine bağlı tek bir popülasyondan ayrılarak farklı bölgelerde evrildiği konusunda ikiye ayrılmıştır.
Afrika'dan çıkış

Chris Stringer ve Peter Andrews tarafından geliştirilen Afrika'dan çıkış modeline göre Homo sapiens 200.000 yıl önce Afrika'da evrildi. 70.000 ile 50.000 yıl önce Homo sapiens Afrika'dan göç etmeye başladı ve sonunda Avrupa ve Asya'daki diğer Homo türleriyle yer değiştirdi. Afrika'dan çıkış modeli kadın mitokondriyal DNA (mtDNA) ile erkek Y kromozomunun kullanıldığı araştırmalar tarafından desteklenir. MtDNA'nın 133 farklı çeşidinin kullanılarak inşa edildiği jenealojik soy ağaçlarının analizlerinden sonra araştırmacılar, günümüzde yaşayan tüm insanların Mitokondriyal Havva adı verilen Afrikalı bir kadın atadan türemiş olduğu sonucuna varmıştır. Bunun yanında Afrika'dan çıkış modeli, insanlardaki mitokondriyal genetik çeşitliliğin Afrika nüfusu içinde en yüksek olduğu bulgusu tarafından da desteklenmektedir.
Afrika'dan tek bir çıkışla mı yoksa birden çok çıkışlarla mı göç olup olmadığı konusunda farklı teoriler vardır. Afrika'dan çoklu çıkışlarla göç etme ve dağılma modeli son yıllarda genetik, dilsel ve arkeolojik kanıtların desteğini kazanmış olan Güney yayılım kuramını içerir. Bu teoriye göre modern insan, 70.000 yıl önce Afrika Boynuzu'ndan başlayarak Asya kıtasının güney kıyıları boyunca yol alıp yayılmaya başlamıştır. Bu grup, Doğu Akdeniz kıyılarına kıyasla çok daha erken bir dönemde, ilk olarak Güneydoğu Asya ve Okyanusya bölgelerine yayılmış olup tarih öncesi bu ilk yerleşimler bu bölgelerde erken dönem insanlarına ait arkeolojik sitelerden elde edilen bulgular tarafından da desteklenmektedir. İkinci bir göç dalgası ise Sina Yarımadası üzerinden Asya'ya doğru olmuş ve insan kitlelerinin geniş Avrasya topraklarına yoğun şekilde yerleşmesiyle sonuçlanmıştır. Bu ikinci grup daha gelişmiş aletlere ve teknolojiye sahip olup ilk gruba kıyasla kıyısal besin kaynaklarından ve deniz ürünlerinden de daha az bağımlı idi. Diğer ilk grubun dağılımlarına dair birçok kanıt, her buzul çağın sonunda yükselen deniz seviyesi nedeniyle tahrip olmuştur. Çoklu göçlerle dağılım modeli, Avrasya, Güneydoğu Asya ve Okyanusya nüfusunun hepsinin aynı mitokondriyal DNA soylarından geldiğini gösteren, bu yüzden Afrika'dan tek bir göç olduğunu ve bu göçün Afrika dışında yaşayan tüm insan nüfusunu ortaya çıkardığını kanıtlayan çalışmalarla çelişmektedir.
Sarah Tishkoff'un başkanlığında Afrika'daki genetik çeşitliliğe dair yapılan geniş çaplı bir çalışma, San halkının Afrika genelinde 121 farklı etnik grup arasında en büyük genetik çeşitliliğe sahip olduğunu ve bu halkın 14 farklı "atalara ilişkin nüfus kümeleri"nden birisi olduğunu göstermiştir. Bu araştırma aynı zamanda modern insanın göçünün kökenini de Namibya ve Angola kıyı sınırı yakınlarındaki Güney Batı Afrika olarak tespit etmiştir.
Bazı antropolog ve arkeologlar tarafından da öne sürülen Toba felaket kuramına göre, yaklaşık 70.000 yıl önce Endonezya'nın Sumatra adasındaki Toba Gölü'nün büyük çapta bir patlaması küresel sonuçlara yol açarak o dönemde yaşayan insanların çoğunun ölümüne ve günümüz insanlarının genetik mirasına etki edecek şekilde bir genetik darboğaza neden olmuştur.
Çok bölgeli evrim modeli
Çok bölgeli evrim, Milford H. Wolpoff tarafından 1988 yılında insan evriminin örüntü ve şekillerini açıklamak için önerilmiş olan bir modeldir. Çok bölgeli evrim modeli, insan evriminin 2,5 milyon yıl öncesi Pleistosen dönemden günümüze kadar kesintisiz olarak sadece tek bir insan türünde gerçekleştiğini ve insanın dünya genelinde Homo erectus'tan modern Homo sapiens'e evrilmesiyle meydana geldiği görüşüne dayanır. Çoklu bölge hipotezine göre, fosiller ve genetik veriler insanın dünya çapında gerçekleşen evrimine dair kanıtlar olup Afrika'dan çıkış modeli tarafından öne sürülen insanın sadece Afrika kökenli olduğu ve Afrika'daki atalardan evrildiği varsayımıyla çelişmektedir. Nitekim Richard Leakey, bu tartışmayı çözmek için fosil kanıtların yetersiz olduğu görüşündedir. Ancak son zamanlarda özellikle Y-kromozom DNA'sı ve mitokondriyal DNA'daki haplo gruplara dair yapılan çalışmalar insanın Afrika kökenli olduğunu büyük ölçüde desteklemektedir. Bunun yanında otozomal DNA'dan elde edilen kanıtlar da ağırlıklı olarak insanın Afrika kökenli olduğunu desteklemektedir. Aynı zamanda modern insanda ilksel ve arkaik karışımların da olduğu bazı çalışmalar tarafından gösterilmiştir. Yakın zamanda Neandertal genomu sıralaması ile Denisova insanı genom sıralaması da bu karışımların meydana geldiğini gösterebilmiştir. Buna göre Afrika kıtası dışında yaşayan modern insanlar genomlarında %2-4 arası Neandertal alelleri taşırken bazı Melanezyalılar buna ek olarak %4-6 arasında Denisova alelleri de taşımaktadır. Bu yeni sonuçlar, mutlak yorumlanması dışında Afrika'dan çıkış modeli ile de çelişmemektedir. Toba volkan felaketi nedeniyle oluşan genetik darboğaz atlatıldıktan sonra oldukça küçük bir grup Afrika'dan ayrıldıktan sonra -büyük olasılıkla Orta Doğu'da, hatta Kuzey Afrika'da henüz tam olarak ayrılmadan önce- Neandertallerle melezleştiler ve onların hala ağırlıklı olarak Afrikalı olan torunları dünyanın tüm bölgelerine yayıldılar. Onların torunlarından bir kısmı da -muhtemelen Güneydoğu Asya'da- Denisova insanlarıyla karışarak Melanezya'ya yerleştiler. Bunun yanında Neandertal ve Denisova insanının majör histokompatibilite kompleksi haplotipleri (MHC sınıf I molekülleri) aynı zamanda modern Avrasya ve Okyanusya nüfuslarında da tespit edilmiştir.
Bilinen insan evrimi araştırmacıları

|


Türler listesi
|
Hominin türleri ile cinslerini içeren liste soldan sağa ve yukarıdan aşağı doğru olmak üzere kronolojik sıraya göre sıralanmıştır.
|
Ayrıca bakınız
- İnsan evriminin fosil listesi
- Fosil primatlar listesi
- Piltdown Adamı
- Arkeogenetik
- İkili kalıtım teorisi
- Ataların Hikayesi
Yaşamın evrimsel tarihi
- Abiyogenez
- Endosimbiyoz teorisi
- Hayvanların evrimi
- Kalbin evrimi
- Sinir sistemlerinin evrimi
- Gözün evrimi
- Kordalıların evrimi
- Omurgalıların evrimi
- Balıkların evrimi
- Sudan karaya geçiş
- Memelilerin evrimi
- Primatların kökeni ve evrimi
- Maymunların evrimi
- Hominoidlerin evrimi
- Gibon-insan son ortak ata
- Orangutan-insan son ortak ata
Dış bağlantılar
- [12] İncredible Human Journey, Tr. Altyazılı belgesel, Dr. Alice Roberts
- [13]23 Ekim 2014 tarihinde Wayback Machine sitesinde arşivlendi. İnsanın evrimsel yolculuğu, belgesel, Türkçe dublaj
- [www.youtube.com/watch?v=XlrjNEMPSQs] Kökenlerimiz-1, Prof.Dr.Alice Roberts
- [www.youtube.com/watch?v=OrjM47L_hnE] Kökenlerimiz-2, Prof.Dr.Alice Roberts
- [www.youtube.com/watch?v=Qus01tIocdQ] Kökenlerimiz-3, Prof.Dr.Alice Roberts
- "La odisea de la especie - Bölüm 1". Belgesel (İspanyolca). 28 Ocak 2016 tarihinde kaynağından arşivlendi. Erişim tarihi: 27 Kasım 2010.
- "La odisea de la especie - Bölüm 2". Belgesel (İspanyolca). 28 Ocak 2016 tarihinde kaynağından arşivlendi. Erişim tarihi: 27 Kasım 2010.
- "La odisea de la especie - Bölüm 3". Belgesel (İspanyolca). 28 Ocak 2016 tarihinde kaynağından arşivlendi. Erişim tarihi: 27 Kasım 2010.
- Şenel, Alaeddin. Kemirgenlerden Sömürgenlere İNSANLIK TARİHİ. Ankara: İmge Kitabevi Yayınları. ss. 1107 sayfa. ISBN 975-533-483-1.
| Alt konular | |
|---|---|
| Bölgelere göre genetik tarih | |